Plumeria Resources
Plumeria Ruba
Related Images:
Branching Inducers on Plumeria
Branching Inducers on Plumeria
To produce more branches per cut from a pruned plumeria limb. UH experiments suggest that lanolin pastes of cytokinins can improve the number of buds which break following pruning. It is desirable to increase lateral shoot production after pruning for cultural control both in commercial flower production and landscapes as well as for plants grown in containers (Kwon and Criley, 1991).
Procedure:
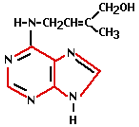
Apply 6-benzylaminopurine (BA) in a lanolin paste to pruned tips. Kwon and Criley (1991) used single stemmed 2 year old plants of P. rubra in 35 x 22 cm pots containing a medium of equal parts soil, peat, and perlite. The plants were decapitated at 30 cm above the soil line 3 weeks after repotting. Various concentrations of the growth regulator solution was then applied to be absorbed into the cut surface. We will use 2 mg/g and 4 mg/g (BA/lanolin) solutions (2000 and 4000ppm). Four plants should be used for each treatment plus 4 control plants. Data to collect are; (a)number of days to bud break for each plant, (b)number of shoots initiated and (c)surviving and (d)the length of these at 4 months after treatment.
The following passage is quoted from a letter from Dr. Richard A. Criley, dated September 16, 1994. It outlines where BA and lanolin may be obtained, how to mix them, how to apply the paste to plumeria, and addresses the question of the shelf life of the mixture.
“The application of chemical branch inducing substances in lanolin would be an interesting set of studies for PSA members to try. You can get plain old lanolin at many pharmacies or order it through chemical supply houses. It comes as a yellow, very sticky fatty substance. You can melt it in a double boiler and dissolve or suspend substances in it. My recommendation is to weigh out the stuff in the same container you want to melt it in – then you are sure of concentrations.
The benzyladenine is not water soluble to any great extent. I usually dissolve it in a small amount of acetone or alcohol or DMSO. You can also use 0.5 N HCL (small amount) to dissolve it, then dilute in water and add to the melted lanolin. Stir well to make a uniform mixture. Allow to cool. I use it by smearing a glob on the cut stump of a plumeria branch using a finger covered with a latex glove finger. Rates to try: 2 or 4 mg BA in 1 gram of lanolin. I have a mixture from 4 – 5 years ago that still has activity.
You can also pour the BA-lanolin paste mix while still liquid into plastic film canisters. This might be one way to share it around easily. (Maybe you could charge a few dollars per unit and recover the costs.)
The N-6-benzyladenine (BA) is also known as 6-benzylaminopurine.”
Summary:
- Use 6-benzylaminopurine (BA) in a lanolin paste.
- Apply paste to pruned tips. (1) 2 mg BA per gram lanolin. (2) 4 mg BA per gram lanolin.
- These experiments would need control plants which receive a no N-6-benzyladenine . Both the experimental subject and control plant would have to be of the same cultivar to avoid inconsistencies in growth habit and flowering.
References:
Eunoh Kwon and Richard A. Criley (1991), Cytokinin and Ethephon Induce Greater Branching of Pruned Plumeria, Horticulture Digest; Hawaii Cooperative Extension Service, No. 93, March 1991, p. 6-8.
6-benzylaminopurine from Sigma Chemical Company; catalog # B 3408: 1 g, $10.65 + shipping. Lanolin from Sigma Chemical Company; catalog # L 7387: 1 kg, $32.80 + shipping.
Related Images:
The influence of colors on plants
By D. Kroeze MSc., CANNA Research
The primary colors
Researchers have traditionally distinguished seven colors. They are known as the colors of the rainbow, red, orange, yellow, green, blue, indigo and violet, often referred to as the primary and secondary colors. When put together, these primary colors create white light and only become visible when a ray of sunshine is split by a prism.
All objects can absorb the colors within the light, reflect them or allow them to pass through. Science tells us that an object usually adopts the color it isn’t able to absorb itself. Because plants cannot absorb the color green, they must themselves be green. There are those who claim that light measurements show that plants themselves emit light particles in the dark and actively help create their own color. Just like we do, plants sense the light and colors surrounding them, but in a different way!
Red light
Plants are sensitive to the color red in the light spectrum, a sensitivity that arises from the plant having what is called a red light photoreceptor. The receptor is a blue-green pigment called a phytochrome present in the cells of a plant. You could compare phytochrome with an eye that only senses red light.
Red light impacts a plant in many ways. Plants that are grown in plenty of red light are often large, but in general also tall with many branches. If the photoreceptor picks up a large quantity of natural red light, for example in summer when there’s plenty of natural red light, production of a plant hormone (metatopolin) is increased. This hormone prevents the chlorophyll in the plant from being broken down, so that it stays green in spring and summer; advantageously, this is exactly the time that the plant needs its chlorophyll to convert energy coming from the sun into sugar. Red light also influences a plant’s flowering and seed formation.

A prism refracts light into the primary colors
By comparing the quantity of one frequency of red light to the amount of another, far-red, present in the light, makes the plant decide whether to start flowering or not. The non-flowering period can be extended by exposing the plant to red-containing light during the dark period. This will, as a result, also extend the period of time before harvesting, which, of course, most growers would like to prevent. This also explains why it is a bad idea to enter the growing area when it’s dark for any time, even for a quick peek! The red color in light also influences flavor because it increases the concentration of special oils in plants.
Blue light
Plants see blue light as well as red light, using a photoreceptor that is called a cryptochrome. If there is plenty of blue light, as is the case in nature during autumn and winter, this receptor slows down the effect of a hormone called Auxin. This hormone is responsible for the plant’s stem and root growth. Auxin is also responsible for what is referred to as ‘apical dominance’, the phenomenon which causes the central stem to be dominant over side stems. A branch off a main stem would be dominant over its’ own side branches so it inhibits the development of axillary flowers. This causes the plant to create more side stems when exposed to bluish light and the plant stays shorter. This helps us to understand why plants exposed to bluish light are often short and bushy in appearance with a more robust structure.
Experiments with blue light resulted in plants that are wider than usual. This can be explained by the fact that more branches sprout because of diminished apical dominance.

Plants use the quantity of blue light to determine how far to open their stomas. The more blue light, the wider they open their stomas, resulting in an acceleration of their metabolism. High levels of blue light will increase metabolism, and as a consequence accelerate plant growth and development.
Blue light is also responsible for leaves growing towards the light. Blue light avoids the multiplication of leaves around the fruits. A shortage of blue light in the spectrum will quickly cause you to lose 20% of your harvest. Although opinions about this seem to differ, the optimum red to blue light ratio should be 5:1 in general.
Green light and the other colors
Plants are hardly sensitive to green light. As far as we know, they lack receptors for this color. This is probably the case because in practice plants do not absorb this color. Plants which are grown in green light only will be exceedingly weak and rarely grow old.
Clearly, plants only seem to sense those colors for which they have specific receptors. This is based on energy levels provided by each color. Plants are not blind but, up to a certain level, they are color blind. The way plants react to orange and yellow light is quite similar to the reaction on red light. This also accounts for indigo and violet which in reaction are similar to reactions on blue light.
‘Invisible’ light

The plant on the left is taller because it received less red light in relation to far-red light. The plant on the right was exposed to the usual red/far-red light relationship.
Although plants are a bit color blind, they do sense colors, more appropriately energy levels, that are entirely invisible to us humans. For example, plants can perceive far-red light. Plants often utilize the red to far-red relationship. A seed uses this relationship to determine whether to germinate or not. Plants also use that relationship to determine the number of other plants in the immediate surroundings.
Because plants absorb large amounts of red light whilst reflecting far-red light, there will be less red light present in a plant’s immediate surrounding if other plants are in the area too. Seeds will hold off germinating and the plants that are already in place will grow faster in order to emerge above the other plants to acquire sufficient light for their photosynthesis.
The fact that far-red light has the exact the opposite effect to that of red light makes it unsuitable as a light for growing. The traditional light bulb is a rich source of far-red light.
Ultra violet light (UV) also influences plants. Like is the case with blue light, plants perceive this color using the cryptochrome photoreceptor. It is unclear whether other photoreceptors can perceive UV light.
When increasing the quantity of UV light, the concentration of a purplish substance called Anthocyanin goes up. Anthocyanins protect plants against UV radiation, but also against micro-organisms trying to get in. The formation of Anthocyanin can often be seen where flaws occur such as a lack of oxygen. UV light not only damages the plant’s DNA and membranes, but immediately disrupts the process of photosynthesis. Therefore an excess of UV light is unhealthy for plants as well.

Left and right: far-red light is reflected by plants and can thus be used to determine the presence of other plants in the immediate vicinity. Plants can perceive far-red light.
Light is seeing
As we have seen, light is not just essential for plants when it comes to supplying energy for photosynthesis. Although this article only reveals a small part of everything there is to know about light, the colors in it and the effects they have on plants, we did see how plants use colors to regulate many of their processes. Plants are capable of perceiving those colors that matter to them. Those colors give the plant an indication of its general environment and its chances of survival and reproduction.
If your plants are to develop, grow and flower well, the composition of the light is at least as important as its quantity. Do not forget that a plant perceives the composition of both direct and indirect light. Indirect light here refers to the light that is reflected onto a plant by other objects such as walls or other plants.
Related Images:
Florigen, the flowering hormone

Many plants are ‘short-day’ plants, meaning that the plant starts to bloom once the days shorten. This is why horticulturists cut the amount of light from eighteen hours a day to twelve when they want to start the flowering phase. It’s actually a shame from the plant’s perspective: six fewer hours of light a day is six fewer hours of photosynthesis, and thus less energy for your plants in the form of sugars. There is also a substance that can get your plants blooming without your having to cut down on their light.
D. Kroeze, CANNA Research
What really happens when the days get shorter or when you cut back to 12 hours a day? When the light goes down to 12 hours or less, the leaves start to manufacture a substance that triggers flowering, which gets transported to all over the plant. This substance is called florigen or flowering hormone.
The term ‘short-day plant’ isn’t completely ac- curate. It’s not the fact that the days are getting shorter that makes the plant decide to flower, but that the nights are getting longer. Although the difference may seem trivial, it does explain why a night-time visit to your growing space will delay flowering for your plants. When you turn on the light, the plant’s night is over; it has now become too short to stimulate flowering. The plant has to start over counting the hours of darkness from zero.

Other short-day plants include maize, chrysanthemum and chicory. There are also long-day and day-neutral plants.
Examples of long-day plants are spinach, let- tuce and barley. One day-neutral plant is tobacco.
Julius and the discovery of florigen

In 1865 a German scientist named Julius von Sachs discovered that when he transferred sap from a flowering plant to a non-flowering plant, the non-flowering plant started to flower as well. This even happened when the two plants were from different species. Unfortunately, no matter what he tried, he never succeeded in isolating the substance responsible for flowering.
Many after him also have tried in vain to isolate florigen, which made it into something of a mystery. It got to a point where the question was not only what the substance actually was, but whether it even existed – at least until a few years ago. Now, one of the greatest mysteries of plant biology seems to have been solved.
Julius von Sachs (pictured top-left) made other major discoveries besides the existence of florigen. For example, he discovered chloroplasts and the fact that they produce sugar. He also discovered that glucose is stored in the form of starch in granules. In other words, the mystery of florigen was as old as much of the fundamental knowledge in plant biology.

Chloroplasts
The information superhighway
During the quest for florigen, it became clear that the sap flowing through the phloem (vessels) of the plant contained more than water and the sugars produced by photosynthesis in the leaves. As it turned out, many semiochemicals (substances that send signals to the plant) are dissolved in the phloem sap. These are mostly small molecules in very low concentrations. The phloem transports information from one place in the plant to another, including the signal to flower, in the form of these substances. This is why the phloem is also known as ‘the information superhighway.
Actually, florigen had been found a few years earlier, but its function had not been discovered until recently. You may wonder why it took so long to find florigen. Here’s the reason: Once the night length has crossed a certain threshold, the leaves produce a signal to start manufacturing florigen. The substance is only made in the growing points of the plant; a different substance, which reacts with the substance from the leaves, occurs only in the cells of growing points. The two substances together are actually the real florigen.
In addition, but no less important, is the fact that these are very tiny molecules, which were only discovered in the last few years. Until then, laboratory equipment was simply not advanced enough.
 The phloem (red) is the living vascular tissue of the plant, through which mainly sugars and water are transported from the top down. Besides the phloem there is also xylem (pink), dead tissue that transports nutrients and water up from the roots.
The phloem (red) is the living vascular tissue of the plant, through which mainly sugars and water are transported from the top down. Besides the phloem there is also xylem (pink), dead tissue that transports nutrients and water up from the roots.
The future is smiling?
After more than 140 years, the quest for florigen is finally over. A great mystery has been solved. This is fine for science, but what does it mean for your average person? The answer is easy: a lot! Manipulating florigen has enormous potential.
Its discovery will bring about a revolution, in particular for conventional agriculture. Greenhouse horticulture will see increased yields from more hours of light. Scientists are especially thinking about growing crops in places where it was previously impossible, such as growing some tropical crops in Northern Europe. But a lot may change for tropical regions as well. The shortened growth time will mean that more crops can be grown in one growing season than is possible now.
Besides this immediate effect on food production, there will also be a revolution for seed companies. For example, fruit trees could be made to flower in the first year and so can be crossed with each other within months instead of the usual years breeders spend waiting for the first flowers.
For hobbyists, it will of course be the higher yields that make the applications of florigen most interesting.
We should not get carried away. It will be many years before we can make practical use of florigen. One thing is certain: its discovery will change agriculture and plant breeding forever.
From Gentech to hunger
Why the question mark after ‘the future is smiling?’ because florigen can’t simply be added to a plant. Biotechnology companies will have to provide crops with the information they need to make florigen themselves independent of day length, using genetic modification. Since this gene will initially be put into one or two crop varieties, these few varieties will quickly drive out local varieties (genetic erosion).
This will mainly be a problem for developing countries where agricultural production is now less than optimum, and where these new crops could greatly improve food production. At first, these monocultures of just a few varieties on such a huge scale will produce lots of food, but in the long term will lead to enormous problems from diseases. And what are you going to eat when the food crops are gone?
That’s the next challenge!
Related Images:
Plant hormones
Plant hormones
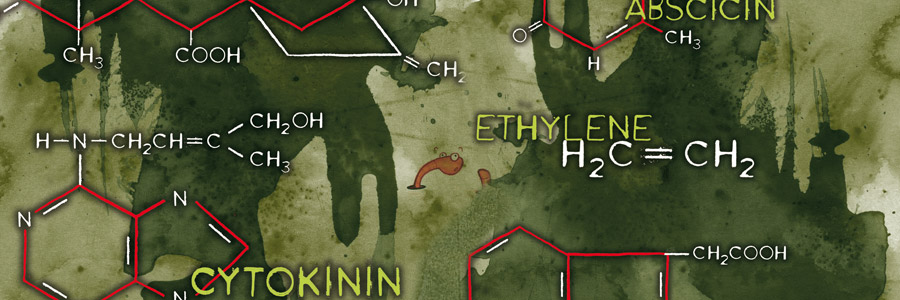
Hormones are organic molecules that can influence the physiology and development of plants and animals even at low concentrations. Hormones play an important role in the growth and flowering of the plant and many other things. This article briefly explains how plant hormones work in plants and how hormones ensure that plants flower.
Hormones are produced by and transported throughout the entire plant. Simply put, they are signals, chemical signals, that can be sent and received throughout the entire plant. A leaf can and will transmit a signal to the end of the stem telling it to form flowers for example. The most well-known plant hormones are auxin, gibberellin, cytokinin, ethylene and abscisin (abscisic acid). In addition, it has been demonstrated that brassino-steriods, salicylates and jasmonates also function in a similar way to hormones. Hormones can also occur bonded to sugars or amino acids. In this form they are inactive and provide storage. The hormones can be released again and become active under various conditions such as the influence of gravity or light for example.
Auxin
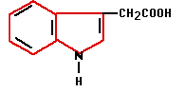 In the 1880’s Charles Darwin and his son Francis started experiments that finally confirmed the existence of plant hormones. They experimented with oats and the influence of light on the direction of growth. Auxin was the plant hormone whose processes were demonstrated during these experiments. Auxin is produced in the plant’s growing points both above the ground and in the roots. Auxin influences water absorption, cell division and cell stretching (it softens cell walls) among other things. Because auxin promotes the formation of roots on stems it is used in a variety of forms in rooting hormones.
In the 1880’s Charles Darwin and his son Francis started experiments that finally confirmed the existence of plant hormones. They experimented with oats and the influence of light on the direction of growth. Auxin was the plant hormone whose processes were demonstrated during these experiments. Auxin is produced in the plant’s growing points both above the ground and in the roots. Auxin influences water absorption, cell division and cell stretching (it softens cell walls) among other things. Because auxin promotes the formation of roots on stems it is used in a variety of forms in rooting hormones.
Experiments carried out by CANNA have shown that the effect of administering auxin depends very much on the concentration and method of application used for each plant type. With weak concentrations flower formation is stimulated slightly and ripening takes longer. With high concentrations there is an inhibiting effect on growth accompanied by deformities and tumour-like symptoms.Auxin that is produced in the tops of plants is capable of inhibiting the development of side shoots. This symptom is known as apical dominance. Removing the main tip stops the inhibiting effect and side shoots then develop which will eventually result in a broader plant. Where crop spacing allows only a few plants per square meter it is worthwhile removing the main tip as this makes it possible to use the light more efficiently. It’s also necessary to remove the tips regularly to achieve a good stock plant for propagation so that it will grow many more side shoots.
Gibberellin
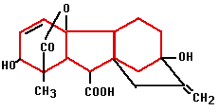 Gibberellin was first isolated in 1935 in Japan by Yabuta. The gibberellin was acquired from a fungus that had been the cause of reduced productivity for Japanese rice farmers for centuries. The gibberellin initially gave better growth but later in the season it caused sterile fruits. Generally speaking, gibberellins work as growth accelerators because of cell stretching and cell division. They ensure that seeds germinate and flowers form in plants that need long days. Gibberellin is often used in the cultivation of fruit to help unfertilized pears and apples develop fully.
Gibberellin was first isolated in 1935 in Japan by Yabuta. The gibberellin was acquired from a fungus that had been the cause of reduced productivity for Japanese rice farmers for centuries. The gibberellin initially gave better growth but later in the season it caused sterile fruits. Generally speaking, gibberellins work as growth accelerators because of cell stretching and cell division. They ensure that seeds germinate and flowers form in plants that need long days. Gibberellin is often used in the cultivation of fruit to help unfertilized pears and apples develop fully.
Administering gibberellin to short-day plants, or autumn flowerers, as they are also known, very quickly gives clear effects even at low concentrations. Plants become light green in colour and stems split open because of the fast growth (photo 1). The plant’s speed of growth can reach 10 cm per day! Administering gibberellin during the vegetative phase causes plants to start flowering more slowly. Gibberellin is for short-day plants as testosterone is for people. It stimulates the formation of typically male organs and longer plants; longer internodes and male flowers in dioecious plants. When the pollen from these flowers is used to fertilize female flowers, seeds are created that always produce female plants.

Photo 1: Stem that has torn open because of growing too fast after gibberellin was administered.
Certain environmental influences can also cause the production of extra gibberellin. Plants will make more gibberellin in poorly lighted conditions, which causes them to become long and look lanky. Another effect is seen when the lamp is too close to the plant. Buds that are flowering can start to shoot again if the lamp is too close. This will cause the tops to become long and thin. To prevent this, the distance from the plant to the lamp during flower formation must be at least 50 cm for a 600W lamp.
Cytokinin
 Cytokinin activity was first demonstrated in 1913. 30 years later it was discovered that a natural substance present in coconut milk was capable of helping plant cells multiply. Cytokinin was the responsible hormone for this. Cytokinin is known as the hormone responsible for cell division. It stimulates the metabolism and the formation of flowers on side shoots and as such is a counterpart to auxin. The cytokinin concentration is highest in young organs (e.g. seeds, fruits, young leaves and root tips). High concentrations in an organ or tissue will stimulate the transport of sugars to those tissues or organs.
Cytokinin activity was first demonstrated in 1913. 30 years later it was discovered that a natural substance present in coconut milk was capable of helping plant cells multiply. Cytokinin was the responsible hormone for this. Cytokinin is known as the hormone responsible for cell division. It stimulates the metabolism and the formation of flowers on side shoots and as such is a counterpart to auxin. The cytokinin concentration is highest in young organs (e.g. seeds, fruits, young leaves and root tips). High concentrations in an organ or tissue will stimulate the transport of sugars to those tissues or organs.
Administering cytokinin leads to greater leaf surface area and faster flower formation. However, the time that flowering finishes is comparable to untreated plants. Cytokinin can be seen as a counterpart to gibberellin in this regard because it stimulates the formation of female flowers on male plants.
Ethylene
 The practical use of ethylene comes from the time of Old Egypt when figs were scored to make them ripen faster. In 1934 it was discovered that plants produce ethylene themselves, which enables them to regulate fruit ripening. Ethylene is the least complex plant hormone from the molecular point of view and is produced by all organs. It is a gaseous hormone which is transported via the spaces between plant cells. It is responsible for fruit ripening, inhibition of growth, and leaf abscission (shedding). Ethylene has a stimulating effect on flower formation with certain types of plants (i.e. pineapples, mangoes and lychees). Administering ethylene results in smaller plants and flowering finishes a lot quicker. The flowers ripen too quickly and consequently remain small.
The practical use of ethylene comes from the time of Old Egypt when figs were scored to make them ripen faster. In 1934 it was discovered that plants produce ethylene themselves, which enables them to regulate fruit ripening. Ethylene is the least complex plant hormone from the molecular point of view and is produced by all organs. It is a gaseous hormone which is transported via the spaces between plant cells. It is responsible for fruit ripening, inhibition of growth, and leaf abscission (shedding). Ethylene has a stimulating effect on flower formation with certain types of plants (i.e. pineapples, mangoes and lychees). Administering ethylene results in smaller plants and flowering finishes a lot quicker. The flowers ripen too quickly and consequently remain small.
Because plants can be very sensitive to ethylene, the concentration is expressed in parts per billion parts of air (ppb). Concentrations of just 10 ppb can cause abnormalities in tomatoes. In situations where ripening flowers come in contact with young plants there is the risk of accelerated ripening in the young plants. The ethylene that is produced can reach the young plants via the air. Ventilating occasionally (once per day) will remove the ethylene that has formed. High concentrations cause leaves to turn yellow immediately.
Ethylene can also accumulate around roots if they are wet for too long. This can lead to leaf chlorosis, stem thickening, leaves bending towards the stem and greater susceptibility to diseases.
In stress situations, for example when there is disease present or damage to the plant, the plant produces more ethylene, which causes it to remain smaller and finish flowering faster. Mechanical stress such as air movement can also cause the plants to produce extra ethylene, which will result in smaller plants with thick, sturdier stems. When the fans are too close to the plants there will be too much stress and this will adversely affect the yield.
Abscisin
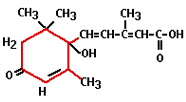 Abscisin was first isolated in 1963 and has the Latin word abscissio (breaking off) to thank for its name. This is because people thought that abscisin was responsible for the breaking off (shedding) of leaves and fruits, however, it was later shown that ethylene plays a much more direct role in this.
Abscisin was first isolated in 1963 and has the Latin word abscissio (breaking off) to thank for its name. This is because people thought that abscisin was responsible for the breaking off (shedding) of leaves and fruits, however, it was later shown that ethylene plays a much more direct role in this.
Abscisin is produced in the chloroplasts of older leaves and has both inhibiting (growth) and stimulating (protein storage) characteristics. When there is a large supply of abscisin to the growing points of the stem and roots, cell division stops and the plant enters a rest period.
Abscisin is an important hormone as far as stress situations are concerned. It is responsible for closing the stomata when the plant is under water stress due to continuing high temperatures, low atmospheric humidity or an EC in the feeding medium that is too high.
Flower formation in short-day plants
Even though a lot of research has already been done into the changeover from growth to flowering in plants, it still hasn’t been explained how this mechanism works exactly. In the case of short-day plants, the formation and development of flowers depends on the precise length of the night. Short-day plants will flower when the night-time period is longer than 12 hours. It is important that it is really dark during this period because the plant is only capable of measuring the period of darkness and not the period of light. Almost any light level during the dark phase will affect the cycle. This is measured in the leaves, which then send a signal to the furthermost ends of the branches instructing them to form flowers. The hormone that gives this signal is called florigen. So it is theoretically possible, for example, to use material from flowering plants to stimulate other plants to flower under 18 hours of light.
Different hormones play an important role in the phase following the first setting of the flower buds. So cytokinin and auxin play an important role in the further formation and growth of the flowers. Abscisin and ethylene are important during ripening.
Using hormone preparations
If you want to experiment with plant hormone preparations, pay close attention to how, when and how much hormone you use. The final effect depends on many factors such as the time of administering (which phase, time of the day), the route chosen for administering (leaf or roots) and the concentration. The final effect of administering hormones can depend very much on the concentration used. For instance: weak concentrations of Auxin stimulate root growth while strong concentrations cause extra ethylene production, which, in turn, causes the plant to finish flowering faster.